inTRO to ipRGCs ( Intrinsically Photosensitive Retinal Ganglion Cells)
Rods, cones, and…ipRGCs?
For almost a century and a half, it was believed that the mammalian retina only had two types of photoreceptors, rods and cones. That assumption was not proven to be false until some studies in the late 1990s proved the existence of a third kind mammalian photoreceptor that differed greatly from rods and cones. These new types of photoreceptors are actually Retinal Ganglion Cells (RGCs) that are intrinsically photosensitive (ipRGCs) or in other words, naturally sensitive to light.
Though the official evidence to determine that ipRGCs actually existed did not come to fruition until the late 1990’s, this third class of photoreceptor had already been hypothesized nearly seven decades prior in 1927 by a graduate student named Clyde Keeler. During one of his studies, he observed mice that lacked nearly all rod and cone function as a result of severe retinal degradation and left them functionally blind. Keeler noticed that regardless of the lack in rods and cones, the mice still had a very strong and significant pupillary constriction in response to light, and he determined that this response must have been the result of some third photoreceptor in the retina. The lack of concrete evidence for a whole new photoreceptor at the time resulted in this pupillary response being explained away by other scientists. However, in 1999, Russell Foster and his team would finally provide the concrete evidence that Keeler needed in order to prove the existence of the third photoreceptor. ← First hypothesis of another receptor
Foster et al. observed a similar situation as Keelers, but in their case, the mice being observed were genetically engineered to not have any rods or cones. Regardless of the missing rods and cones, the rats still displayed strong pupillary light reflexes and were even able to shift their circadian rhythms accordingly with light exposure schedules. With these studies complete, the presence of a third photoreceptor was almost confirmed, but many still weren’t convinced because nobody had found another light-sensitive molecule (opsin) in the mammalian retina yet. ← First proof of another receptor
The discovery of melanopsin in photosensitive skin cells of frogs occurred in 1998 and in the following four years, studies determined that the very same opsin was being expressed in a small amount of RGCs in both mouse and human retinas. This discovery allowed scientists to easily mark ipRGCs and confirm their existence which finally put to rest the debate of whether or not there was a third class of photoreceptor.
So they exist but, what do they do?
ipRGCs differ greatly from rods and cones when it comes to how they work. Their main function in the body is to signal the intensity of ambient light levels (irradiance) to the brain. These signals are largely used for non-image-forming visual reflexes that are subconscious, such as pupillary constriction, neuroendocrine regulation, and synchronizing daily (“circadian”) physiological rhythms to the light it responds to. This means that the way they respond to light is also quite different from rods and cones.
As I mentioned before, these photoreceptors use melanopsin as their photopigment and that makes cells more responsive to light at around 480nm (blue light). In the graph below, you can see that this wavelength is significantly different (≥20 nm) from the best wavelengths for stimulating rods and cones.
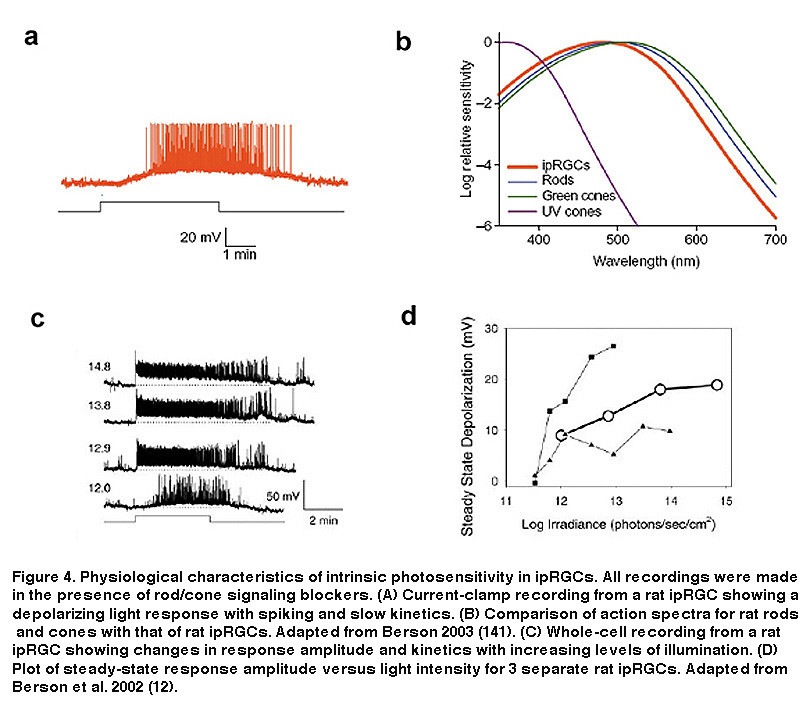
When compared to rods and cones, ipRGCs are much less sensitive to light, respond to light much more slowly, have far lower spatial resolution, and also have the unique ability to directly communicate with higher visual centers of the brain.
Although ipRGCs function as photoreceptors, it was found that they nevertheless receive synaptic input from the circuits of rods and cones. This means that ipRGCs have both an intrinsic light response coming from melanopsin and an extrinsic one that is mediated by synaptic input from rods and cones. The light response caused by melanopsin is markedly different from that of rods and cones, but in both cases any light will continuously be signalled for at least 10 hours. Basically, some intraretinal wires are crossed which cause ipRGCs to have both an intrinsic and sluggish light response as well as an extrinsic, rod/cone driven, rapid photoresponse. There is an ongoing debate about the significance of this extrinsic synaptic input and the role rods and cones play in determining our circadian rhythms. All things considered, we know that ipRGCs as a whole are responsible for many of the subconscious responses we have to light and our current methods of modulating those subconscious light responses is to alter total irradiance or control the amount of blue light.
An interesting case:
In a research article by Mouland et al. the team determined whether the effective light intensity reported by melanopsin (blue light ~480nm) was a more important determinant of the circadian impacts caused by light than that of cones. The ability to determine mealopsin’s contribution to circadian light responses comes from the evolution of a color science technique which is referred to with multiple names, such as receptor silent substitution or metamerism in colorimetry. Metamerism occurs when two colors appear to match under a specific lighting condition.
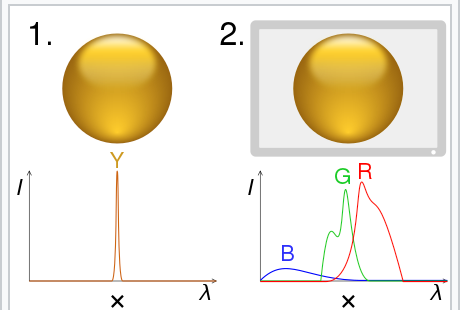
This technique allows for the stimulation of specific photoreceptor classes, like ipRGCs, which have previously been shown to increase melatonin suppression and alertness. Mouland and his colleagues determined the circadian impacts of the photoreceptors by recording electrophysiological activity from the suprachiasmatic nucleus (SCN) of anaesthetised mice while they were presented with movies. The movies were either high or low contrast and had varying irradiances specialized for the distinct photoreceptor classes.
During the experiment, the energy response recorded from the SCN was nearly identical to that coming from melanopsin when melanopsin activity was at its highest. This means that at the most effective melanopic irradiance, cone-derived signals had little effect on SCN firing rates. In general, steps in melanopic irradiance were determined to be the most significant factor accounting for light-induced changes in SCN activity. The only significant effects that were recorded by changes in cone-directed irradiance occurred in low contrast movie conditions. Basically, this study suggests that cones do have an impact on the circadian signal going to the SCN in some conditions but the influence of melanopsin on the circadian signal was overall more significant because it was consistently present across all conditions.
This blog post was written by Arcascope’s intern, Ali Abdalla. Thanks, Ali!
This post used Webvision as a major resource. Thanks to Dustin Graham and Kwoon Wong for the excellent review.